
BRINTON AND TOWNSEND: EUPHAUSIIDS
IN
THE GULF OF CALIFORNIA-THE
1957
CRUISES
CalCOFI
Rep.,
Vol.
XXI,
1980
EUPHAUSIIDS IN THE GULF OF CALIFORNIA-THE
1957
CRUISES
E.
BRINTON AND A.W. TOWNSEND
Marine Life Research Group
Scripps institution
of Oceanography
La
Jolla. CA
92093
ABSTRACT
Euphasiid crustaceans in the Gulf of California were
examined from four bimonthly CalCOFI grid cruises
during February through August of
1957.
Of the nine
species found to regularly inhabit the Gulf,
Nematoscelis
dificilis
and
Nyctiphanes simplex
are common to the
warm-temperate California Current. These have the
broadest ranges in the Gulf, peaking in abundance and
reproducing maximally during February-April and Feb-
ruary-June respectively, before intense August heating
takes place in the
Gulf.
Euphausia eximia,
a species
having high densities at zones considered marginal to the
eastern tropical Pacific, also varies little in range during
the year, consistently occupying the southern half of the
Gulf. Tropical
Nematoscelis gracilis
shows a range com-
plementary
to
that
of
N. dificilis;
these species overlap in
the southern Gulf. Three
Euphausia
species of the tropi-
cal Pacific occupy the southern Gulf in February-April,
expanding northward during June-August but, like the
cool-water species, scarcely reproducing in the Gulf dur-
ing the warm season. The distributions and abundances of
the species and their life stages, particularly the youngest
larvae, are described in relation to seasonal variation in
flow and temperature in the Gulf.
RESUMEN
Se examinaron cruskiceos eufausidos del Golfo de
California tornados durante cuatro cruceros bimensuales
de CalCOFI entre febrero y agosto de
1957.
De las nueve
especies encontradas que regularmente habitan en el
Golfo,
Nematoscelis dificilis
y
Nyctiphanes simplex
son comunes en las aguas templada-calidas de la Co-
rriente de California. Estas especies presentan una dis-
tribucion amplia en el Golfo, alcanzando una maxima
abundancia y reproduccion durante febrero-abril
y
febre-
ro-junio, respectivamente, antes de llegar el calor intenso
de agosto.
Euphausia eximia,
una especie de altas den-
sidades en las zonas consideradas marginales a1 Pacifico
tropical oriental, tambien varia poco en
su
amplitud de
distribucion durante el aiio, ocupando consistentemente
la mitad sur
del
Golfo.
Nematoscelis gracilis,
es tropical
y
muestra una distribucion complementaria con
N.
d$-
ficilis;
estas especies concurren en el sur del Golfo. Tres
especies de
Euphausia
del Pacifico tropical ocupan el
sur del Golfo en febrero-abril, extendiendose hacia el
norte durante junio-agosto, pero, como las especieg de
agua fria, escasamente se reproducen en el Golfo durante
la estacion calida. Las distribuciones
y
abundancias de
IManuxript
reccivcd
14
April
I960
I
las especies y de
sus
etapas de vida, en particular las de
las larvas mas juveniles, se describen en relacion con las
variaciones estacionales en el flujo y las temperaturas de
las aguas del Golfo.
INTRODUCTION
The Gulf of California is inhabited by dense stocks of
plankton (Osario-Tafall
1946;
Zeitzschel
1969).
This
appendix of the North Pacific Ocean communicates.
with the 'eastern boundary circulation at the northern limit
of the eastern tropical Pacific, which is characterized by
its distinctive oxygen-deficient layer. The
1
000-km
axis
of the Gulf extends from the mouth at the tropic, 23"27'N,
to latitude 32"N, which is within the belt of the warm-
temperate Californian Transition Zone, discussed most
recently by Newman
(1979).
The Gulf parallels the
terminal part of the California Current, which meanders
southward on the other side of the Baja California
peninsula.
Little attention has been paid to ways in which the
Gulfs nearly enclosed planktonic populations range along
this
stretch of water, which is recognized on the basis of
the fish fauna to be one of temperate-tropical transition
(Walker
1960).
Indeed, only sparse information exists as
to the zooplankton species presently occupying the Gulf
or to their distribution. Data are best for Foraminifera
(Bradshaw
1959;
Parker
1973),
Chaetognatha, Siphono-
phora, and Scyphomedusae (Alvariiio
1969),
and pon-
tellid Copepoda (Fleminger
1967, 1975).
Hyperiid Am-
phipoda are presently under study by Siegel-Causey,
whose first findings (in press) presage description of a
diverse amphipod fauna. Existing knowledge of the plank-
ton of the Gulf of California has been reviewed by Brinton
et al. (in press).
Euphausiids sampled across the mouth of the Gulf
(Brinton
1979)
were grouped as
1)
residual from the main
stream of the California Current
to
the north (e.g.
Nem-
atoscelis dificilis, Euphausia pacixca,
and
Thysano-
essa gregaria),
2)
species proliferafing at the productive
margins of the eastern tropical Pacific (e.g.
Euphausia
eximia
and the associated, more coastal
Nyctiphanes
simplex),
3)
endemic equatorial vertical migrators adap
ted to the oxygen-dificient regions
(Nematoscelis gra-
cilis,
Euphausia diomedeae,
and indistinctly associated
E. distinguenda
and
E. lamelligera),
and
4)
non- or short-
distance migrating warm-water cosmopolites that avoid
the oxygen deficiency (e.g.
Stylocheiron afine,
S.
carina tum
1.
The
Gulf
deepens into basins with depths up to
1,000
m
21
1

BRINTON AND TOWNSEND: EUPHAUSIIDS
IN
THE GULF
OF
CALIFORNIA-THE
1957
CRUISES
CalCOFI Rep.,
Vol.
XXI,
1980
in the north and 3,500 m in the south. Hence, there is
sufficient vertical space for occupancy by the
Euphausia
and
Nematoscelis
species which undergo vertical migra-
tions
of
300 or 400 m. The extent to which euphausiids
penetrate the Gulf had been known previously only on the
basis of a September-October 1952 CalCOFI cruise
along the southernmost part
of
the western side (Brinton
1962; these 1952 distributions are detailed here along
with the 1957 distributions).
Half of the area
of
the Gulf is neritic, with the broadest
shelf
in
the northern end and along the eastern side
(Figure 1). Although most euphausiids have oceanic
rather than neritic ranges, nevertheless there is a species
with neritic affinities (Brinton 1962; Figures 113, 114)
corresponding to each, but one, of the Pacific's ten
littoral provinces recognized by Ekman (1953). Two of
these occur in the Gulf.
Nyctiphanes simplex's
range
corresponds with Ekman's American Temperate Tropi-
cal Zone and
Euphausia lamelligera's
with the Pacific
Tropical American.
Ships
of
the National Marine Fisheries Service and the
Scripps Institution of Oceanography occupied a stan-
dardized grid of stations in the Gulf during 1956 (three
cruises) and 1957 (four cruises). All employed the Cal-
COFI techniques regularly used at that time for samp
ling zooplankton; that is, oblique tows to 140 m, using a
1-m
net
of
0.55-mm-mesh width, following Ahlstrom
(1 948). The 1957 cruises provided the most seasonal and
geographical coverage and were selected for this study.
Net tows generally strained 300 to 400 m3 of water and
were standardized to 1,000 m3. Samples were aliquoted
and counted using methods described in Brinton (1979).
A biogeographical analysis of fish eggs and larvae in
the Gulf (Moser et al. 1974) was based on samples from
three of the four cruises discussed here and from three
1956 cruises. Hence, the 1957 fish distributions can be
strictly compared with these euphausiid distributions.
Interest in examining biology
of
Gulf plankton in rela-
tion to oceanographic processes has heightened due, in
part, to the importance to Mexico of understanding regen-
erative cycles in these productive coastal waters and also
to emerging awareness among oceanographers of the
Gulfs distinctive topography, its unique climate, and the
substantial containment
of
its biota. Ayala-Castanares
(1979) has characterized it as a natural laboratory.
THE
PHYSICAL
ENVIRONMENT
Upwelling
and
Mixing
The climate and circulation of Gulf waters are in-
fluenced by seasonally reversing winds and changes in
incident heat. Upwelling, determined by the divergence of
the Ekman transport, is expected to be strongest along the
eastern side of the
Gulf
during the northeasterly winds
SAN
FELIPE
BAH~A
DE
ADAIR
BAH~
k.
-.
'0.
"
"
:10
Figure
1.
The
Gulf
of
California. Edge of shelf and
maximum
depths
of
principal
basins are indicated.
of winter and spring and then to shift to the western side
during southwesterly winds of summer and early fall
(Roden 1971). This
is
illustrated by the temperature
distributions in Roden and Groves (1959) and in Figures
2
and 3 of the present paper; these distributions are based
on the 1957 cruise measurements.
The many islands, particularly along the western Gulf,
provide much extra coastline which augments the extent
of mixing processes and upwelling:
1)
in channels be-
tween islands or between islands and the coast, tidal mix-
212

BRINTON AND TOWNSEND: EUPHAUSIIDS
IN
THE GULF
OF
CALIFORNIA-THE
1957
CRUISES
CalCOFI Rep.,
Vol.
XXI,
1980
.j
.":b;
'&
cy.
Flgure 2. Temperature at 10-m
depth
during the four 1957
cruises and during Septem-
ber-October 1952 (after
Anonymous 1963).
ing can be as intense as it is over the northern shelf and
2)
island topography may be optimal for persistent upwell-
ing because there is always some coastline normal to the
shifting wind.. Such diverse features preclude generalized
explanations of nutrient renewal, plankton production,
and hence, the distribution of plankton in the Gulf.
Temperature
at
10-m Depth (Figure
2)
These temperatures are representative of the mixed
layer into which the older life stages
of
most vertically
migrating euphausiid species occurring here usually as-
cend in the nighttime. Larvae live only in this layer and
the upper part
of
the thermocline (Brinton
1979).
February.
Isotherms were roughly zonal, ranging from
14°C
in the island channels to
20-2
1
"C
at the mouth
of
the
Gulf.
The "island channels" (cf. Regional Differ-
ences in Population Structure, below) refer to waters sur-
rounding Islas Angel de la Guarda, Salsipuedes, San
Lorenzo, San Esteban, San Pedro Martir, and Tiburon in
the zone
28?4-29?4"N.
April.
Isotherms assumed meridional alignment. Tem-
peratures of
15"-17"C
prevailed around Isla Angel de la
Guarda, southward to the west of Isla Tiburon. This cool
area was surrounded by the
18"-20"
water which ex-
tended over most of the Gulf. At mid-Gulf,
26-27"N,
temperatures were above
2
1
"C
along the western side but
were less than
20°C
to the east. The western side of the
southern Gulf was
22-23°C
and the eastern,
17°C.
Thus,
the eastern side was cooler in the southern Gulf than far-
ther north.
June.
The advance of seasonal warming continued,
reaching
23-26°C
over most of the Gulf. Only in the
mixed waters of the northwestern island channels were
temperatures as low as
21-22°C.
Along the western side
of the mid- and southern Gulf, temperatures responded to
effects of the southerly winds and were
2
to
3°C
cooler
than to the east, thus differing from the condition in April.
August.
Temperatures of
29-3
1
"C
now prevailed al-
most throughout the Gulf, with lowest values, to
28"C,
again only in the northwestern island channels.
September-October.
Temperatures observed in
1952
in the southern Gulf agreed with those of August
1957,
indicating that summer conditions may persist into
October.
213

BFUNTON AND TOWNSEND: EUPHAUSIIDS
IN
THE GULF OF CALIFORNIA-THE
1957
CRUISES
CalCOFI
Rep.,
Vol.
XXI,
1980
Figure
3
Temperature at 100-m depth (data from Reid et al
1965)
Positions of hydrographic casts are indicated as small dots Stations for temperature
profiles
in
Figure
4
are indicated by large dots
Temperature at
100-m
Depth (Figure
3)
At 100-m depth, temperature varies seasonally in the
Gulf, whereas at 200 m, change is insignificant (Figure
4). At the deep mouth of the Gulf, 100-m temperature re-
mained nearly constant, 14-16"C, throughout the year,
whereas in the northern, shallower half
of
the Gulf,
water at this depth warmed from 13-14°C in February-
April to 19-23°C in August. In the sector of the island
channels (Zone
B,
Figures 4, 18), this variation reflected
the appearance of a summer thermocline following in-
tense winter and spring mixing through the water column.
In the mid- and southern Gulf the summer increase was
evidently caused both by convection and downward con-
ductive transfer from a surface subjected to progressively
intense heating.
Seasonal variation at 100
m
was greatest in the eastern
Gulf (Figures 3, 4), evidently due to more penetrating
June-August warming than took place along the western
side where upwelling had become relatively stronger.
One hundred meters approximates the deep limit of the
nighttime level of most euphausiids considered here (ex-
cept the somewhat deeper living adults of
Nematoscelis
dzficilis)
and is near the upper limit of the levels to which
migrating juveniles descend in the daytime, whereas adults
of most species go deeper.
February.
Upward bending
of
subsurface isotherms
was conspicuous toward the eastern side of the Gulf. This
was not evident in the 10-m temperature distribution,
indicating that upwelled water was not reaching the
surface layer.
April.
At mid-Gulf (26-28"N) there was
<
14°C water
on both sides, but the eastern half was still generally a
little cooler than the western.
June.
The April situation was now reversed, as also in
the surface layer. Isotherms now emerged toward the
west.
August.
The progression of summer warming was con-
spicuous, particularly over the broad shelf of the northern
Gulf and along the eastern side. Ridging of subsurface
isotherms along the Gulfs axis was evident toward the
south.
Oxygen
The oxygen-deficient layer in the eastern tropical Pa-
cific lies between the thermocline and about 700 m. The
[02]
in this layer may be as low as
0.05
mlhter, which is
near the limit of measurability. Water having this ex-
treme minimum extended from 21
"N
off
Cab0 Corrientes
at the mouth of the Gulf, southward
to
1
1
"N during June
of
1974 (Brinton 1979). During 1957 this layer was
diluted when it entered the Gulf. For example, at
23"N
[02]
of 0.1 mlhiter was unusual at any season and never
occurred in water above 300 m except near the western
side in April (Roden and Groves 1959). Moreover, the
lowest
[02] found half-way up the Gulf, at 27"N, was 0.2
ml/liter at 500-m depth.
214

BRINTON AND TOWNSEND: EUPHAUSIIDS
IN
THE
GULF
OF CALIFORNIA-THE
1957
CRUISES
CalCOFI Rep.,
Vol.
XXI,
1980
TEMPERATURE
("C)
1CP
12'
14O
16'
18'
2P 22' 24' 26' 28"
30"
32"
....I
I
I
I I I I
."I
I
112-1
I
I
I
AUG,I/
100
E
v
I
I-
200
n
300
0
100
200
300
CWEST
Sta.
127.40
DWEST
Sta.
139.40
10"
12' 14' 16'
18'
20"
22"
24' 26'
28'
30"
32"
-
-
-
-
I
Sta.
117.50
-
i
t'
Figure
4.
Temperature profiles for February. April, June, and August of 1957 at localities on eastern and western sides of Gulf of California. Zones
(B,
C
and
D)
and localities are shown in Figure 18 (data
from
Reid et al. 1965).
Oxygen deficiency has much to do
1)
with creating an
environment for endemism among euphausiids in the
eastern tropical Pacific and 2) with sharply restricting
subtropical species and warm-water cosmopolites there.
However, it appears not to be sufficiently shoal, extreme,
or widespread in the Gulf to have had the same domi-
nating role in the selection of which species are present.
Nevertheless, the depth of a given low-[O2] surface
may be used to show the spread within the Gulf of this
salient property
of
eastern equatorial water as it relates to
the 0-300-m range
of
depth in which most euphausiids are
expected to live. In the basins
of
the northern Gulf, the
1.5
ml/liter [02] surface is below 200-300 m during all
seasons (Figure
5).
It
is
deepest during April-June in the
strongly mixed waters
of
the western island channels. By
mid-Gulf it shoals
to
100-200
m, and at the mouth, to
70-
100
m. This surface tilts upward toward the east during
the northeasterly winds
of
February-April, then upward
toward the west in June.
Least [02] within the 0-300 m layer is at localities
where the
1.5
ml/liter surface shoals to
70
m
or
less.
Roden and Groves showed that low values of
0.1-0.5
215

BRINTON AND TOWNSEND: EUPHAUSIIDS
IN
THE GULF
OF
CALIFORNIA-THE
1957
CRUISES
CalCOFI Rep.,
Vol.
XXI,
1980
Figure
5.
Depth of
1.5
ml/liter concentration of oxygen (data from Reid et al.
1965)
ml/liter are at
100-300
m in the southern Gulf, with the
upper limit
of
the 0.5-mVliter layer shoalest in February
(40-80
m) and deepest in August
(150-250
m).
Salinity
Seawater evaporation exceeds freshwater runoff into
the Gulf (Roden and Groves
1959),
but surface salinity
beyond embayments ranges between
34.4
and
35.9
a/oo.
This compares with amaximum salinity
of
about
34.0
'/,,
in the open sea to the west (Love
1973).
Therefore
salinity per se is not expected to influence the distri-
bution and abundance
of
plankton in the Gulf. Estuaries
and lagoons along the southeastern coast may neverthe-
less contribute nutrients to the Gulf, as indicated by the
primary productivity maxima, which were observed sea-
ward of that coast during August of
1972,
associated with
relatively low surface salinities of
34.4-34.5"/,,
(Gilmar-
tin and Revelante
1978).
of the Gulf during February and April. Flow at
200
m-
the daytime depths of many migrating euphausiids-was
then generally consistent in direction with surface flow.
According
to
Roden and Groves this upper level outflow
is compensated by inflow at greater depths.
By June, surface flow had reversed, entering the Gulf
across most of the mouth and persisting toward the north
as a stream which shifted toward the eastern side by
August. Flow at
200
m
was opposite
to
surface flow
during the summer.
Direct observations supplemented by computations of
geostrophic flow have been made of cyclonic cells of
circulation extending across the width of the Gulf during
August
1978 (Emilsson
and Alatorre in press). Such a
cell in the 0-50-m layer in the southern Gulf provided
inflow along the eastern shelf and outflow near the tip of
Baja California not unlike that described for August
1957.
A similar eddy farther inside the Gulf was also
Currents
The geostrophic flow diagrammed in Figure
6
(after
Wyllie
1966)
shows mean surface flow to have been out
indicated. This is pertinent in discussing exchange be-
tween the Gulf and the adjacent Pacific and how plank-
tonic stocks may be maintained in the Gulf.
216

BRINTON AND TOWNSEND: EUPHAUSIIDS IN THE GULF
OF
CALIFORNIA-THE 1957 CRUISES
CalCOFI
Rep.,
Vol.
XXI,
1980
APPROXIMATE
CURRENT
SPEED
GEOSTROPHIC
FLOW
AT
200m
DEPTH
(RELATIVE
TO
500
DECIBARS)
AWROXIMATE
CURRENT
SPEED
Figure
6
Geostrophic flow at surface and
200
m,
relative
to
500
decibar surface, inferred from casts
at
indlcated localities (after Wyllie
1966)
217

BRINTON AND TOWNSEND: EUPHAUSIIDS
IN
THE GULF OF CALIFORNIA-THE
1957
CRUISES
CalCOFI
Rep.,
Vol.
XXI,
1980
A&t@hunes
shplex
1-49
per
1000m3
50-499
500-4,999
5,000-49,999
0
5
50.000
Figure
7.
The coastal neritic species,
Nyctiphanes
simplex.
THE
SPECIES
DISTRIBUTIONS
Nyctiphanes
simplex
(Figure
7)
This euphausiid has the strongest neritic ties of those
inhabiting the subtropical Californian region. Its range
typically extends along Baja California northward to
southern California, 34”N. During the warm year 1958,
it reached 40”N
off
northern California.’
N.
simplex
is
the most abundant and widespread euphausiid in the Gulf
of California during all seasons and has been observed in
dense swarms by day (Steinbeck and Ricketts 1941) and
in fields of luminescence at night (authors’ personal
ob-
servation), as
“Nyctiphanes”
(night shining) implies.
The vertical range of
N.
simplex
seems to vary with
‘Published reports
on
the distributions of the Gulf species outside of the area of this study
will
not
be
cited repeatedly in the text
to
follow. These references are,
for
the California Current, Brinton
(1967a,
b,
1973): forthe eastern tropxal Pacific and its vansition with the California Current at
the
mouth
ofthe Gulf, Brinton
(1
979); for the overall Pacific, Brinton (1962); and for the global
picture, Mauchline and Fisher
(1969)
and Brinton (1975).
locality (Brinton 1967a, 1979). Clearest evidence of
diurnal migration has been found seaward of the shelf.
Species
of
Nyctiphanes
in other oceans have been de-
scribed as descending close to the shelf in the daytime
where they may feed in the superficial sediments (Mauch-
line and Fisher 1969; Gros and Cochard 1978)..High
densities (>5,000/1,000 m3) in the Gulf were found in
both day and night samplings.
During 1957, maximum numbers were generally near-
shore
or
over the shelf. In February, when surface iso-
therms were oriented in an east-west direction (Figure 2),
there were three zones with high densities, 27-28”N, 25-
26”N, and 24-24?4”N on the eastern side. These were
located alternately with zones in which densities were mod-
erate, generally <500/1,000 m3. The February distribu-
tion extended to the southernmost part of the Gulf, assoc-
iated with southerly flow in the upper levels (Figure 6).
218

BRINTON AND TOWNSEND: EUPHAUSIIDS IN THE GULF OF CALIFORNIA-THE
1957
CRUISES
CalCOFI Rep.,
Voi.
XXI,
1980
..
Nemutoscehs
oM%c/as
1-49
per
1000m3
50-499
500-4.999
5.000-49,999
140-
100.
Figure
8.
The warm-temperate species, Nemafoscefis difficifis.
In April, high densities were along the Gulfs margins,
particularly the eastern side. These extended as far south
as the mouth, 24"N, where surface temperatures of 17-
20°C persisted, now due
to
upwelling. Except for low
numbers at the mouth, densities of
N.
simplex
were
>500/1,000 m3 almost everywhere in the Gulf, as the
overall population peaked.
By June the upper water masses
of
the Gulf were
warmer by 3-6°C than in April, and mean flow in the
southern half of the Gulf was northerly. Highest densities
were then general to the northern Gulf but were infre-
quent south of the island channels, diminishing to zero at
the mouth.
When temperatures increased to 28-3
1
"C in August,
the population became reduced. The overall range of
N.
simplex
remained unchanged except for retraction from
the northernmost shelf region. Highest densities were
now around the island channels where temperatures were
lowest, to 28°C. Peak numbers had been outside
of
these
channels during the previous months. Pockets
of
moder-
ate density were present on the western side of the Gulf
to
24"S, evidently associated with summer mixing processes
along that coast. The 1952 data on
N.
simplex
suggest
that those processes persist into October.
Nematoscelis difficilis
(Figure
8)
In the oceanic North Pacific
Nematoscelis dificilis
inhabits the temperate-subtropical transition zone of the
North Pacific Drift, 40-45" N
(=
Subarctic-Central in
water mass terms). It is also prominent throughout the
California Current from southern Oregon to Baja Cali-
fornia but
is
known from the Gulf of California on
the basis
of
three low-density records near Bahia Conceg
cion, September-October 1952 (Figure
8). This popula-
219

BiUNTON AND TOWNSEND: EUPHAUSIIDS
IN
THE GULF OF CALIFORNIA-THE
1957
CRUISES
CalCOFI Rep.,
Vol.
XXI,
1980
Nemtoscehs
qruc,%s
1-49
per
i000m3
@@
50-499
40'
200
P
209
40-
140'
100'
Figure
9.
The tropical
Nernatoscelis
species,
N. gracilis
tion was interpreted (Brinton 1962) as relict-isolated by
tropical water at the mouth
of
the Gulf. However, the
range
of
N. dzficilis
in the Gulf during 1957 nearly
overlapped that of
Nyctiphanes simplex,
and this species
was second in abundance of the Gulf euphausiids. These
were altogether unexpected findings.
It is safe to state that
N.
dzflcilis
lives deeper than
Nyctiphanes simplex,
though their vertical ranges over-
lap. The profiles for those species to the west of Cali-
fornia and Baja California showed
N.
simplex
tending to
stay above approximately 100 m and adult
N. difJicilis
below the mixed layer, except where stratification had
broken down, as with upwelling (Brinton 1967a, 1979).
(Weibe and D'Abramo
[
19721 showed that most
Nema-
toscelis megalops,
a close sibling and geographical ana-
logue in the Atlantic
of
N.
dzficilis,
are below 100 m at
night in the Mediterranean Sea. Wiebe and Boyd
[
19781
showed that
N. megalops
was spread through the upper
600 to
800
m in Gulf Stream cold core rings and
apparently did not engage in diurnal vertical migrations.)
The February 1957 distribution showed
N.
dificilis
concentrated toward the western side of the Gulf. High
densities extended to the mouth. Though February data
were lacking for the northern Gulf, this month is seen
to
be generally optimal for
N. dzficilis.
Penetration into the
southeastern Gulf
of
the 20°C isotherm at 10 m is associ-
ated with fewest numbers, but the presence
ofN.
dzzcilis
this far south assures exchange with the California Cur-
rent population (cf. February 1957 distribution to west of
Baja California in Brinton 1967b).
In April, instances of >5,000/1,000 m3 were fewer
than in February in the southern half of the Gulf but did
occur along both coasts to about 26"s.
From June to August, the diminishing population
220

BFUNTON AND TOWNSEND: EUPHAUSIIDS IN THE GULF OF CALIFORNIA-THE
1957
CRUISES
CalCOFI Rep.,
Vol.
XXI,
1980
10.
Euphausia
eximia,
Euphuusu
mhiu
1-49
per
1000m3
@#
50-499
500-4,999
5,000-49,999
which proliferates at margins of the eastern
tended to concentrate toward the west where coolest
temperatures prevailed. Heated shelf waters to the north
and east evidently provided limited refuge there, with
occurrences primarily in or near the northern island chan-
nels and in the eastern zone south of Bahia Guaymas, 27-
28"N.
Tropical water entering the mouth of the Gulf-
particularly to the east-appears to have compressed the
distribution toward the north and west. Larvae, which
customarily occur in the upper layer now warmed to 28-
30°C, were represented by a single specimen, a furcilia,
in the August catches.
If the vertical range of adult
N.
diflcilis
in the Gulf is
greater than 200 m, as was supposed to be the case
off
southern California, this might induce mortality when the
euphausiids are carried into neritic waters. However, in
the terminal part of the California Current
off
Baja Cali-
foria, 23"N, adults migrated through less than 200 m,
400
20.
0.
20'
40'
140.
100-
tropical Pacific.
apparently limited to strata where
[02]
was greater than
about
1.5
ml/liter. Such a vertical range in the Gulf could
help to explain the extensive presence
of
N.
dzflcilis
shoreward
of
the basins.
During June and August in the mid- and southern
sectors of the Gulf (Figure 3,4), high temperatures in the
upper 100 m would be expected to limit
N.
dzBciZis
to
below that depth, and shoaling
of
the 1.5-mUliter oxygen
surface to less than
100
m could further narrow the verti-
cal range, thereby all but eliminating this organism from
the southern Gulf by August.
Nematoscelis gracilis
(Figure
9)
This species belongs to the euphausiid assemblage of
the eastern tropical Pacific, while also ranging along the
equatorial Indo-Pacific belt. The June 1974 data from the
mouth of the Gulf
of
California (Brinton 1979) showed
22
I
BRINTON AND TOWNSEND: EUPHAUSIIDS
IN
THE GULF OF CALIFORNIA-THE
1957
CRUISES
CalCOFI
Rep.,
Vol.
XXI,
1980
Euphuusiu
dshhquendu
1-49
per
1000m3
50-499
500-4,999
5.000-49,999
Figure
11.
Euphausia distinguenda, endemic
to
the eastern tropical Pacific.
that the northern limit
of
N.
gracilis
abuts the southern
limit
of
congener
N.
dz@cilis
there. Adults of both species
were beneath the 0-40-m layer at night, except at an
up
welling locality to the west of Cab0 San Lucas. In the
daytime
N.
gracilis
was within the extreme
[OZ]
mini-
mum layer, 300-400 m, while
N.
dificilis
was above 200
m, apparently limited by the
1.0-
or 1.5-mlhter
[Oz]
surface. Larvae and juveniles of both species ranged
within or into the mixed layer, day and night.
During February
of
1957,
N.
gracilis
was present only
in the southern Gulf. Its most northern presence was to
the east where the oxygen minimum was shoalest (Figure
5) and where
N.
dzficilis
was sparse. Flow at 200-m
depth shows this distribution
to
be within a system of slow
eddy circulation. Elsewhere, where
N.
dzficilis
was dom-
inant, flow was southerly (Figure
6).
The April distribution extended farthest north on the
western side, as far as Isla Espiritu Santo, 24%"N. This
was within the warmest waters in the Gulf->2l0C at 10
m (Figure 2) and >14"C at 100 m (Figure 3).
June warming and strong northerly flow in the upper
levels (Figure
6)
brought
N.
gracilis
to the northern island
channels, 29"N, though its uninterrupted geographical
distribution extended only to the Guaymas Basin, 27%"N.
By August, northerly transport to mid-Gulf was re-
duced. There, eddy circulation developed, and the range
ofN.
gracilis
retracted southward. The high temperatures
general to the Gulf in August should be suitable for this
species, in keeping with its distribution across the eastern
tropical Pacific. Hence, a southerly trend in flow within
the subsurface habitat of the bulk of
N.
gracilis
may be
limiting.
.
Euphausia eximia
(Figure
10)
E.
eximia
is abundant in the California Current to the
west of Baja California, 21-30"N. The range extends
222

BRINTON AND TOWNSEND: EUPHAUSIIDS
IN
THE GULF OF CALIFORNIA-THE 1957 CRUISES
CalCOFI Rep.,
Vol.
XXI,
1980
Euphuusiu
/ume/hgem
1-49
per
1000m3
50-499
500-4,999
=
5,000-49,999
0
t
50,000
Figure
12.
Euphausia lamelligera, coastal to the eastern tropical Pacific.
across the eastern tropical Pacific, but the species is
sparse, nonreproductive, and avoids the mixed layer in
waters where surface temperature exceeds 26°C and/or
[OZ]
in the minimum layer is <O.lml/liter (Le. low). This
includes the region between the equator and 2
1
ON
at the
mouth of the Gulf.
E.
eximia
in the Gulf shifted little with season, appear-
ing as a resident population from 28"N southward. As
with the two warm-temperate species,
Nemutoscelis dificilis
and
Nyctiphunes simplex,
highest densities of
E.
eximia
were during February. Maxima for
E.
eximia
were in a
region of sluggish circulation in the southern Gulf. The
most northern presence of significant numbers wak in
June when upper level flow was strongly toward the
north, particularly on the western side. Lowest densities
were in August when temperatures >26"C extended to
25-60-m depth (Figure
4).
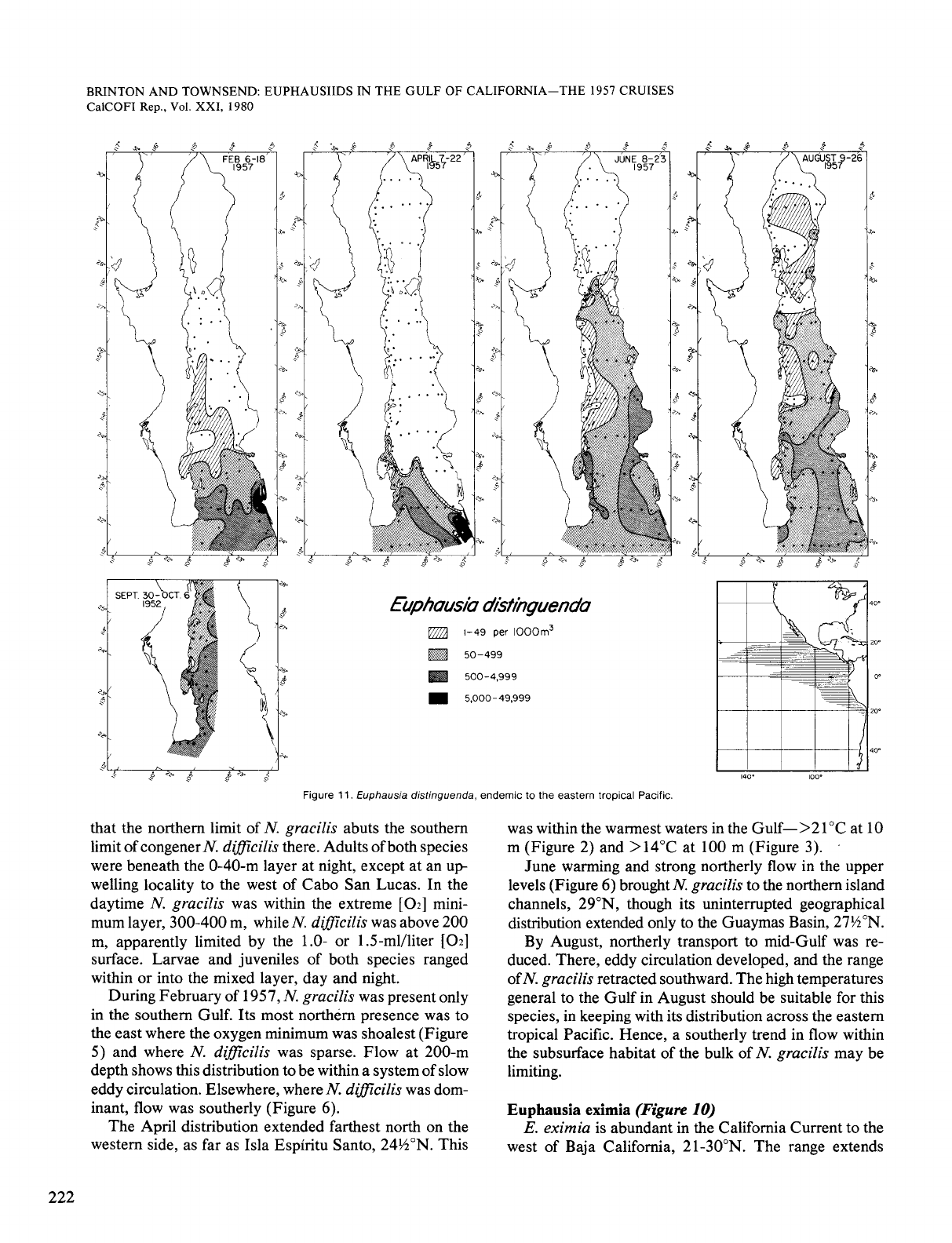
Euphausia distinguenda
(Figure
12)
E.
distinguenda
is endemic to the eastern tropical
Pacific where it is widespread and is the most abundant
euphausiid. The vertical range extends up into the warm-
est layer there and it is also tolerant of lowest
[OZ],
0.05
ml/liter,
at
daytime depths. In both respects,
E.
distin-
guenda
differs from E.
eximia.
It
also
differs from
E.
eximia
in that its distribution in the Gulf fluctuates great-
ly with season.
In February,
E.
distinguendu
was as far north as
27"N
off
Bahia Concepcion. This may be a residual tongue; the
August range showed that
E.
distinguenda
is distributed
nearly throughout the Gulf late in the year. Also, this
mid-Gulf presence is to the west of the southerly flow at
both the surface and 200 m (Figure 6).
As during February, high densities in April were at the
mouth of the Gulf, with maxima over the eastern shelf.
223

BRINTON AND TOWNSEND: EUPHAUSIIDS
IN
THE GULF OF CALIFORNIA-THE
1957
CRUISES
CalCOFI Rep.,
Vol.
XXI,
1980
Euphuusiu
teneru
1-49
per
1000m3
50-499
500-4.999
Figure
13.
Euphausia
tenera,
oceanic in tropical seas.
E.
distinguenda
moved up the Gulf during June-August
when northerly flow developed, mainly on the western
side. Upper level temperatures now exceeded 22°C at 10
m and 15°C at
100
m. Presence of this organism in shelf
waters of the northern Gulf in August must have resulted
from northward flow through the island channels, followed
by branching flow toward the east.
Euphausia lamelligera
(Figure
12)
E.
lamelligera
is more coastal than
E.
distinguenda
in
the eastern tropical Pacific where both are endemic. The
two species are particularly closely related (Brinton 1975,
1979), but their overlapping coastal-versus-oceanic
ranges within the tropics indicate incomplete habitat
separation.
There appears to
be
even less separation in the
Gulf
than farther south. Here there is much shelf and slope area
in relation to basin, and a population having basic afin-
ities with one system could readily be mixed with the
other. Thus, the overlapping of
E.
distinguenda
and
E.
lamilligera
in the Gulf parallels the similarity between
the distributions of the Gulfs two warm temperate euphau-
siids-typically oceanic
Nematoscelis dificilis
and coastal
Nyctiphanes simplex.
Whereas the February-April range
of E.
lamelligera
scarcely reached mid-Gulf, 27"N, the June-August per-
iod
of
northerly flow brought it to the northern shelf,
mainly along the eastern slope. Breaks in continuity
appeared in the zone of the island channels, near 29"N
(cf.
E.
distinguenda).
Highest densities were regularly in
eastern waters at the mouth
of
the Gulf.
Euphausia tenera
(Figure
13)
E.
tenera
has a broadly tropical distribution around the
globe. It is the smallest
Euphausia
species-slender,
224

BRINTON AND TOWNSEND: EUPHAUSIIDS
IN
THE GULF
OF
CALIFORNIA-THE
1957
CRUISES
CalCOFI
Rep.,
Vol.
XXI,
1980
Fiogure
14.
The two Stylocheiron species found in the Gulf
of
California.
S.
affine
excludes the relatively divergent Central Form.
rarely reaching
11
mm in length, compared with
12
and
14
mm
for the more robust
E.
lumelliggera
and
E.
distin-
guenda
or with as much as
25
mm for
E.
eximiu
in the
Gulf. However the diurnal vertical migration ofE.
teneru,
as determined in the eastern tropical Pacific, is
as
exten-
sive
as that of the others, about
0-300
m for the adult.
During February and April,
E.
teneru
was limited to
the southern Gulf. During the June and August periods of
warming and inflow, the distribution extended barely to
mid-Gulf, mainly within the northerly current near the
eastern side. This, together with
E.
teneru’s
consistently
low density over much of the southeastern shelf at the
mouth,
24-26”N,
contributed to this being the
Euphuusia
species at lowest density in the Gulf. These character-
istics attest
to
the midocean, though not necessarily blue-
water, affinities ofE.
feneru
(Brinton
1962, 1979),
rela-
tive to the eastern boundary current affinities of the pre-
dQminant species in the Gulf.
is present here as the East Equatorial Form. The overall range shown for
S.
affine
Stylocheiron
affine
and
S.
carinaturn
(Figure
14)
During
1957,
these euphausiids were few, patchy in
occurrence, and restricted
to
the southern Gulf. They
were not present in the September-October
1952
samples.
The intrusions show
no
sigdcant pattern of seasonal
change.
S.
uBne
is represented in the general area by a “Cali-
fornia Current Form” and an “East Equatorial
Form,”
which have been observed to merge
off
the mouth
of
the
Gulf(Brinton
1962).
These, together with a “West Equa-
torial
Form,” are the basis for the composite range shown
in Figure
14,
inset. There were few adult specimens in the
195
7
material, and no analysis of
“Form”
was attempted.
S.
curinutum
has been described as particularly sparse
off
the mouth of the Gulf and, when present there (Brinton
1962, 1979),
as derived from tropical water to the south.
S.
ufine
is a nonmigrator, living mainly within the
thermocline.
S.
curinutum
appears to be a short-distance
225

BRINTON AND TOWNSEND: EUPHAUSIIDS
IN
THE GULF OF CALIFORNIA-THE
1957
CRUISES
CalCOFI
Rep.,
Vol.
XXI,
1980
migrator, between the mixed layer at night and the ther-
mocline in the day. Thus, both are distributed by upper
level flow, being unable to avail of the deep counter-
currents which often serve the strong vertical migrators in
maintaining range stability.
Nematobrachion flexipes
and
Euphausia diomedeae
Two other euphausiid species (distributions not shown
here) were recorded at or near the mouth
of
the Gulf.
Young of
Nematobrachion jlexipes
were at 23”N, near
the tip
of
the Baja California peninsula in June (one
record) and August (two records). These specimens evi-
dently came from the southernmost part of the California
Current, inasmuch as
N. jlexipes
has been found lacking
in the most oxygendeficient part of the eastern tropical
region to the south.
The virtual absence from the Gulf of the widely tropi-
cal species
Euphausia diomedeae
was unexpected. Single
specimens were found at 23”N, south and southeast of
Cab0 San Lucas, in April and June. In August there were
two records at the mouth of the Gulf and one at 27”N in
the northerly stream on the eastern side.
E. diomedeae
is related to
E.
eximia
(e.g. their larvae
are almost indistinguishable), more
so than to
Euphausia
lamelligera, E. dish’nguenda,
or
E. tenera.
Evidently the
Gulf is habitable by
E. eximia,
which “proliferates at the
margins of the eastern tropical Pacific” (Brinton 1979),
and not by its more tropical relative,
E. diomedeae.
E
U
PH
AU
SI
I
D
BIOMASS
In tropical and subtropical seas, the proportion of eu-
phausiid biomass to total zooplankton biomass (wet dis-
placement volume) retained by our nets varies between
about 3% and
50%
among localities. However, within
oceanographic regions there is less variability. This is
seen in the June 1974 values from three regions of the
eastern Pacific (Brinton 1979):
1)
southernmost Cali-
fornia Current,
7
=
12%, range 7-15%; 2) outer Gulf
(Cab0 San Lucas to Cab0 Corrientes),
j;:
=
8%, range 7-
13%; 3) eastern tropical zone (Acapulco to Galapagos
Islands),
E
=
21%, range 9-49%.
These means
of
euphausiid biomass are higher than the
3 to 7% obtained inside the Gulf during 1957 (Figure
15).
However, an annual mean of the four monthly means
of
total zooplankton biomass in the Gulf in 1957 is close
to 300 cc/l,OOO m3. This compares with the regional
1974 means outside the Gulf of 258,
312,
and 305 cc/
1,000 m3 for regions
I),
2), and 3) above, respectively.
These latter values for total biomass were obtained in
June 1974. The June 1957 mean inside the Gulf agreed
with the mean of the four 1957 cruises. This indicates that
biomass in the Gulf, at least in June, may be of the same
order as that in the subtropical-tropical ocean outside.
However, the June 1974 data outside
of
the Gulf were
500
-
“E
0
0
0
L
400
0
I-
2
W
U
300
a
_I
a
E?
5
D
v)
I
0
m
-
2
200
z
0
+
s
100
a
a
_I
8
50
N
ix
0
%
EUPHAUSIIDS
I%
JUN
%
-
m
AUG
3%
4
3
“E
L
al
a
0
2.3
I
0
Figure
15.
Mean euphausiid biomass shown as a percentage of the mean
total biomass for each
Gulf
of
California cruise in
1957.
obtained using a Bongo net, which differs in design from
the 1-m ring net used in the Gulf in 1957. The Bongo net
has
no
anterior bridle to signal a warning to the plankton.
Bongo nets are proving to be more effective than the 1-m
net in catching euphausiids of >8-mm length; smaller
sizes are caught in equal number (authors’ unpublished
observations from California Current samples). Biomass
of specimens 7-14 mm (which includes juveniles of all
Gulf species
+
adults of all except
Euphausia eximia
and
Nematoscelis difficilis)
is underestimated by about
two times when the 1-m net is used; of specimens 15-20
mm (includes adults of
E.
eximia
and
N. difficilis)
by
about four times. Hence, most Gulf species may have
been substantially undersampled, leading to underestima-
tion of euphausiid biomass by a mean of three times. The
estimates of total numbers of euphausiids are much less
affected by difference in the kind of net than are estimates
of biomass because specimens
>8
mm-mostly adults-
are few compared with young stages.
Comparisons of biomass among the Gulf cruises and
between the Gulf cruises and samplings outside the Gulf
may nevertheless aid in placing the Gulfs plankton in
perspective with that of the adjacent ocean. Euphausiid
biomass in the Gulf was nearly constant at 20 cc/I,OOO
m3, or
0.2
g carbon/m2, during February to June (means
for all stations of each cruise; Figure 15). However, ex-
pressed as a percentage of the total, euphausiids were
most, 7%, in February and June, and down to
5%
in
April when total biomass peaked.
This nearly uniform euphausiid biomass during Feb-
ruary to June resulted from the simultaneous buildup in
226

BRINTON AND TOWNSEND: EUPHAUSIIDS
IN
THE GULF
OF
CALIFORNIA-THE
1957
CRUISES
CalCOFI Rep.,
Vol.
XXI,
1980
q
0
\
\
a
v,
FE0
APR
JUN
AUG
Figure
16.
Biomass of
six
euphausiid species, each determined as a mean of
values
for
all stations
of
each of the Gulf of California cruises in
1957.
Nyctiphanes simplex
and decline from February on-
wards in
Nematoscelis dizcilis
(Figure 16). February
peaks in biomass for
Euphausia eximia
and
E. lamel-
ligera
contributed little to the overall euphausiid biomass
at that time, through certainly being significant in the
southernmost part of the Gulf.
Euphausiids decreased to 3% in August, when the two
bulk species were much diminished.
THE DEVELOPMENTAL PHASES-
ABUNDANCES AND DISTRIBUTIONS
During 195 7 in the Gulf, euphausiid recruitment and
survivorship developed in an interpretable way in the
three species that appear to be resident-those sufficiently
numerous and geographically stable to show some inde-
pendence from the populations outside the Gulf. These
species are
Nyctiphanes simplex, Nematoscelis dificilis,
and
Euphausia eximia.
The tropical species entering the Gulf are centered
either 1) in the eastern tropical Pacific to the south
(Euphausia distinguenda
and
E. lamelligera)
or 2) in
the ocean-wide equatorial belt to the south and west
(Nematoscelis gracilis
and
Euphausia tenera,
and
E.
diomedeae
of
which one specimen was found north of the
mouth, 23" 30").
E. distinguenda
and
E. lamelligera
reproduced substantially in the southern Gulf. Few larvae
of
N. gracilis
and
E. tenera
(see below, Proportions
of
Life Phases), and none of
E. diomedeae,
were found in
the Gulf.
The calyptopis is the youngest larval phase retained by
our nets, and its abundance will be used as a measure
of
recruitment to the populations of the species considered
resident. Only general trends in survivorship and growth
may be derived from the four bimonthly sets of samples.
There is some information on life-span of species of
euphausiids similar to those living in the Gulf. Jorgensen
and Matthews (1 975) gave one year as the probably life-
span
of
Nematoscelis megalops
in a Norwegian fjord.
N.
diflcilis
is very closely related to
N. megalops
and has the
same life-span, or a little longer, based on Brinton and
Wyllie's (1976) data from the California Current. Gros
and Cochard (1978) interpreted
Nyctiphanes couchii,
a
northeastern Atlantic relative of
N. simplex,
as living
approximately two and a half years in the Bay of Biscay.
Euphausia pacifica,
in the California Current
off
south-
em California, lives to one and a half years, according to
Brinton (1976); there is no information on the life his-
tories
of
the tropical
Euphausia
species.
Nyctiphanes simplex
In the Gulf,
Nyctiphanes simplex
produced most ca-
lyptopes in February (Figure 17), the mean being 3,600/
1,000 m3, which accounted for 80% of the total for the
four 1957 cruises. Calyptopes declined in numbers as the
year progressed. However,
N. simplex
was the only spe-
cies to produce these larvae in significant numbers as late
as August. High variances (Appendix I) are associated
with these means, and with those below, in the discussion
of
regional variation in the occurrences of life stages.
However, means will nevertheless be used here as indica-
tions of seasonal and regional trends in abundances.
Furcilia larvae increased during February-June, while
calyptopes decreased, indicating improving larval sur-
vivorship as spring progressed. Youngest
N. simplex
ca-
lyptopes of 1-mm length (Boden 1951) must grow to
oldest furcilia: of
5-
or 6-mm length in about a month (cf.
N.
couchii;
Le Roux 1973)-certainly in less than the
twemonth interval between the Gulf cruises. Hence, the
June peak in furcilia: derives from calyptopes produced
somewhat after the apparent peak in February.
Juvenile
N. simplex,
like the furcilia larvae, were most
numerous during April-June, whereas adults showed a
steady increase during February-June, with scarcely a
decline in August.
Nematoscelis difficilis
Ninety percent of the February-August crop of calyp
topes
of
Nematoscelis dificilis
were produced in Feb-
ruary when the mean density was 900/1000m3 (Figure
17). The coinciding February peak in the older larvae
(
furcilia:) indicates that maximum recruitment was ear-
lier than February. (Gopalakrishnan [1973] found the
larval life of
N. dificilis
to be of the order of 30 days.)
Calyptopes declined during April to June and ceased to
be produced by August. Adults and larvae were both
most numerous in February. Whereas
Nyctiphanes sim-
plex
adults increased through the year, those of
N. difi-
cilis
decreased, though in both species the adult stock
remained nearly constant.
It
appears likely that the nu-
merous larvae of February, or somewhat earlier, led to
the April-June peak in juveniles and then to a substan-
227

BRINTON AND TOWNSEND: EUPHAUSIIDS
IN
THE GULF OF CALIFORNIA-THE
1957
CRUISES
CalCOFI Rep.,
Vol.
XXI,
1980
Figure
17.
Seasonal variation in abundances of life phases of five predominant euphausiids. Values are means of all values, each Gulf
of
California cruise in
1957.
See Appendix
I
for data on Nyctiphanes simplex and Nernatoscelis difficils.
tial stock of young adults in August, clearly the most
stressful period for
N. dificilis.
These adults would be
expected to reproduce during the ensuing winter.
These zones are modified from those proposed by Gilbert
and Allen (1943) that corresponded with differences in
diatom abundances and are like those proposed by Round
Euphausia eximia
This species followed nearly the same pattern as
Ne-
matoscelis dzficilis.
Ninety-five percent of all calyptopis
larvae were observed in February, with their production
decreasing nearly
to
zero by August. In
E.
eximia
there
were relatively more furcilia entering the population as
late as June. Adults were then at a second peak, follow-
ing a February maximum. Adults held their numbers into
August, evidently receiving input from the June juveniles,
whereas the younger phases were much reduced.
Euphausia distiriguenda
Of the tropical species,
E.
distinguenda
showed the
most stability through the year, with furcilia, juveniles,
and adults nearly constant in numbers. However, the
youngest larvae (calyptopes) were significantly present
only in February inside the Gulf. Substantial recruit-
ment from the south may have occurred during the June-
August period of northerly flow.
Euphausia iamelligera
The distribution of
E.
Zamelligera
(Figure 12) shows
that the center
of
distribution
of
this coastal species is at
the mouth of the Gulf and southward. As in the other
species, peak recruitment in the Gulf was in February.
Furcilize steadily declined thereafter. Adults remained
nearly constant in numbers, except for a near absence
in
April at the end
of
the spring period of outflow.
Regional Dwerences in
Population
Structures
The Gulf is considered here to consist of four zones
(Figure 18), which roughly reflect common limits of sea-
sonal shifts
in
zonation of the euphausiids (Figures 7-14).
(1967) for phytoplankton floras. Round’s northern and
southern zones apply to euphausiids. His central zone is
subdivided here to separate the distinctive “island chan-
nels,” 28%-29%”N, from “mid-Gulf.’’
Although scarcely 100
km
apart, the two sides
of
fie
Gulf nevertheless differ with respect to
1)
extent of conti-
nental shelf, hence of neritic mixing through the water col-
umn, and 2) nutrient renewal through upwelling in a given
season. In the northern Gulf, in the zone of island chan-
nels and in the mid-Gulf, mean temperatures (10-m
depth) are somewhat lower
on
the western side through-
out the year (except in mid-Gulf in February; Figure 19).
In
the southern Gulf, the eastern side is cooler during
February-April, and the western side during June-August.
Nyctiphanes simplex
and
Nematoscelis dizcilis,
the
two species that utilize most of the area of the Gulf, were
examined for east-west differences in population charac-
teristics in the four zones.
Nyctiphanes simplex
(Figures 18A, 20A). In the north-
ern Gulf and in the island channels (zones A and
B),
calyptopis larvae were more abundant
on
the eastern side
throughout the year, except in August when the western
island channels became the principal refuge for the pre-
sence of calyptopes.
In the southern half of the Gulf (zones
C
and
D)
calyp
topes were commonest on the eastern side in February,
with the maximum shifting to the west during ApriLAugust.
However, the dominance
of
zones A and
B
in the pro-
duction
of
calyptopes is seen in Figure 20A, in which the
mean value for the eastern Gulf during all of February-
June (four zones combined) is well above that for the
western side.
228

BRINTON AND TOWNSEND: EUPHAUSIIDS
IN
THE GULF
OF
CALIFORNIA-THE
1957
CRUISES
CalCOFI Rep.,
Vol.
XXI,
1980
1
I
I
I
I
I
I
I
I
I
I
Figure
18.
Seasonal changes
1957
in abundances of life phases of a) Nycfiphanes simplex and b)Nematosce/is difficilis on eastern and western sides of Gulf of
California. (See Appendix
I
for data.) Zones are approximated from characteristics of euphausiid distributions: A, northern Gulf;
B,
zone of island channels;
C, mid-Gulf;
D,
southern Gulf
Furcilia larvae averaged a little more numerous to the
west in February and August (Figure 20A), largely be-
cause of higher numbers in the western island channels
than in the eastem, zone B (Figure 1SA). During April-
June the overall maximum moved
to
the east, affected
largely by the distribution in the upper half
of
the Gulf
(zones A and B).
Like calyptopes, juveniles and adults were more con-
centrated
to
the east during February-April. The dif-
ference was more significant in the adults, due 1) in Feb-
ruary
to
high numbers
to
the west in zone C, though most
adults were then in zone
D,
and 2) in April
to
high
numbers
to
the west in zone A, to which zone the maxi-
mum
for
adults had then shifted.
By June the maxima for juveniles and adults had
moved toward the west, with the abundances in the north-
ern Gulf (zone A) most strongly affecting the mean values
shown in Figure
20A.
By August, maxima
for
both
of
these older life phases had shifted back to the eastern side:
however, Figure
1
SA again shows that the easterly shift
only took place in the north, though in both zones
A
and
B.
Nematoscelis diflcilis
(Figures 18B,
20B).
In the
nothern Gulf with its extensive shelf, zone A, most calyp
topes were on the western side in April, with the maxi-
mum shifting to the east in June (Figure 1SB). Somewhat
more furciliae were to the east in zone
A
during both April
and June. Juveniles declined almost equally on both sides
of the northern Gulf as the year progressed. More adults
were
to
the west during April-June, but by August there
was no east-west difference.
In zone B, mixing is to considerable depths in channels
between islands and between islands and the mainland.
To
the west, where the deep Canal de las Ballenas and
Canal de Salsipuedes are generally cooler than surround-
ing waters (Figure
19),
larvae were more abundant
229

BRINTON
AND
TOWNSEND:
EUPHAUSIIDS
IN
THE
GULF
OF CALIFORNIA-THE
1957
CRUISES
CalCOFI Rep., Vol.
XXI,
1980
3"
;
A
(Northern
Gulf)
I I I I
Ll
30"
25"
20"
15°C
!
1
Figure 19. The 1957 10-meter temperatures for eastern and western halves of each zone of the Gulf of California, plotted as means and ranges.
l0,000
1
1,000
1
E
Fl
loo^
E
c
t
0
t
Figure
20
The 1957 seasonal abundances of life phases of the two pre-
dominant warm-temperature species, comparing eastern and western
sides of the Gulf of California
A Nyctiphanes simplex.
B
Nematoscelis
difficils (See Appendix
I
for data)
throughout the year than to the east, off Isla Tiburon.
Juveniles declined progressively through the year
on
both
sides of the zone, while numbers of adults fluctuated.
In zone C, mid-Gulf, there were more calyptopes to the
west than to the east during the reproductive period, Feb-
ruary-April. Juveniles were more numerous in this zone
than to the north, and adults were more consistent in num-
bers through the year than elsewhere.
On
both sides of zone
D,
the southern Gulf, calyptopis
and furcilia larvae steadily decreased to zero as the year
progressed. Juveniles and adults were regularly more
abundant
on
the more oceanic, western side.
Thus, significant east-west differences in
N.
dzEciZis
were few:
1) Calyptopes averaged somewhat higher to the west
during February-April (Figure 20B), a consequence of
higher abundances to the west in all four zones. During
June, they were more abundant to the east in three of the
zones and were equally distributed in the fourth.
2) Furcilia were more abundant to the west during
February, particularly in zones B and C, but were equally
distributed during April-June.
3) Juveniles and adults were about four times more
abundant
on
the western side during June-August, with
the least such difference
in
the shallow northern Gulf,
zone A.
TEMPERATURES AT WHICH
YOUNGEST LARVAE OCCURRED
Temperature at 10-m depth in the Gulf ranged be-
tween 13 and 32°C during February-August
1957.
The
youngest larvae sampled (calyptopes)
of
one or another
euphausiid species were found across the full range of
230

BRINTON AND TOWNSEND: EUPHAUSIIDS
IN
THE GULF OF CALIFORNIA-THE
1957
CRUISES
CalCOFI Rep.,
Vol.
XXI,
1980
Eupbausia
disfihguenda
20
JUNE
AUG
temperature, except the maximum,
>31"C
(Figure
21).
However, it was evident (Figure
17)
that in all species
most reproduction was in the winter.
In
this
discussion, a given
"C
includes all increments
between that
"C
and the next, Le.
"14°C"
includes
14-
14.9"C.
February temperatures were the lowest observed,
13-
20"C,
and in April the range was
1522°C.
General
warming had begun by June.
Nyctiphanes simplex
produced
80%
of its calyptopis
larvae in the mid- and southern Gulf (zones
C,
D)
in
February at localities where temperatures were
17-1
8°C
(Figures
2,
21).
Most of the smaller April crop was
where temperatures were
14, 17,
and
20"C,
with the
highest mean at
14°C.
These low temperatures no longer
prevailed in the upper layers in June when the small
number of calyptopes were found mostly at
23-24"C,
near the lower limit of temperature found for that cruise.
In August most of the relatively small maximum of calyp-
topes were found at 27°C-again the lowest temperature
encountered.
From the lowest to the highest temperatures there is a
declining trend in the percentage of stations at which
calyptopis larvae of
N.
simplex
were found, regardless of
numbers. This trend (Figure
21)
is punctuated by spikes
at
14"C, 23"C,
and
27-28"C,
corresponding to the April,
June, and August maxima in absolute numbers.
In
Nematoscelis diflcilis
the February and April tem-
peratures at which calyptopis larvae peaked were
17-
19°C
(Figure
2 1).
This range was general to mid-Gulf in
February and mainly on the western side in April. The
few larvae produced in June were scattered across the full
available range of temperature of
22-27°C.
Percentages
of stations at which calyptopes were found tended to be
highest toward the lower end of the temperature range.
In the three principal
Euphausia
species living in the
southern half of the Gulf during their main period of
reproduction, February-June, calyptopis larvae were
found concentrated at
20°C
during the February maxi-
mum, again at
20°C
in April in the case of
E. eximia,
at
24-26°C
in June, and at
29-30°C
in August. These were
essentially the prevailing temperatures in the Gulf during
the respective seasons. We know nothing of the extent to
which these larvae might be at somewhat greater depths
as the year progressed,
so
as to occur at more nearly
constant temperatures. However, the simple reduction in
numbers of larvae after February in the cases of
E. dis-
tinguenda
and
E. lamelligera
and after April in E.
eximia
indicates that the warmer months are not conducive to
substantial recruitment in the Gulf.
PROPORTIONS
OF
LIFE PHASES
For each of seven euphausiid species, the annual pre
portion of the total made up by each of the four life phases
sampled here is similar. Catch curves, in which means for
the four cruises are averaged (Figure
22),
show furcilia
larvae to be the most numerous phase and adults the least.
There are fewer calyptopes than furcilia: due, in part, to
the relative duration of these phases.
Data
obtained from
rearing
Nematoscelis diflcilis
(Gopalakrishnan
1973),
Nyctiphanes couchii
(Le
Roux
1973),
and
Euphausia
eximia
(M. Knight personal communication) all indicate
that the calyptopis phase lasts about
10-12
days and fur-
cilia phase about twice as long,
18-24
days. Thus, even
without considering interphase mortality, mean numbers
of calyptopes should be half those of furcilia. However,
calyptopes were found to average about one quarter the
number of furcilia. There is clearly substantial escape-
ment by calyptopes through the 0.55-mm meshes of the
net. (Smallest calyptopes of
Nematoscelis diflcilis
and
-1
AUG
JUNE
03
20"
2
5'
30"
-5
0
LA-AA-Ai
15"
500
ul
bo
0
I
:
FEB-APR
MAX
rn
i4~
n
Eipbausia
eximia
-
-
-
20
-
15"
20" 25"
30"
t-FEB MAX4
-
IPR
MAX
-
2
500
-
0
Z
1X
n
v
I
5"
20"
25"
5000
4000
3000
2000
I000
n
4-J
MAX
"
13"
15"
17"
19"
2Io 23'
25"
27"
29'
3I0C
a?lOrn
2
7
13
lI
26
I8
31
40
34
21
2923
34
26
7
16 60
48
3
No
of
Stohons
Figure
21.
The youngest larvae sampled (calyptopes), all four
1957
cruises of
the Gulf
of
California, plotted as a functlon
of
temperature
(10-m
depth).
-
23
1

BRINTON AND TOWNSEND: EUPHAUSIIDS
IN
THE GULF OF CALIFORNIA-THE 1957 CRUISES
CalCOFI Rep.,
Vol.
XXI, 1980
10'
I
0'
-
O
W
z
m
H
0
u
3
!?
IO'
3
n
v
P
-
"E
0
10
0
0
b
m
a
n
W
5
D(l
z
01
CATCH
CURVES
I
1
IL
CALYPTOPIS
FURCILIA JUVENILE
ADULT
Figure
22.
Abundances
of
life phases, all of the
1957
Gulf
of
California cruises
combined.
Nyctiphanes simplex
are about
1
.O-1.3
mm in length and
about one-half of that in depth or width.)
The juvenile and adult phases in warm-temperate eu-
phausiids are expected to be of the order of six months
each, with some adults living to one and one-half or two
years of age (Brinton
1976;
Gros and Cochard
1978).
The shapes of the curves in Figure
22
reflect a combin-
ation of
1)
recruitment intensity,
2)
duration of life-phase,
and
3)
survivorship.
A
rough mean for the ratio of calypto-
pis:furcilia:juvenile:adult
for all seven species is
1
:4:3:0.5.
Euphausia eximia
has relatively most calyptopes, indi-
cating both high recruitment and high larval mortality in
this
largest of the Gulf species.
E. distinguenda
and
E.
tenera
have proportionately more adults than the other
species, indicating either
a
relatively long adult phase or a
greater influx of adults to the Gulf. The similarities among
the curves suggest similarities in life-span-possibly one
year in this strongly seasonal regime.
Whereas
Nyctiphanes
and
Nematoscelis
incubate their
externally attached eggs for a time,
Euphausia
sheds its
eggs directly into the water. This means that the nauplii
(not retained by our nets) of
Euphausia
swim freely but
those of
NyctQhanes
and
Nematoscelis,
whose eggs
hatch as metanauplii and then develop into calyptopes,
are protected. The difference in rearing strategy does not
lead to a difference, here,
in
the ratio of calyptopes that
survive to the next phase.
SUMMARY AND DISCUSSION
The mouth of the Gulf of California is located almost
exactly on the tropic,
23"27'N.
To the west, this line
approximates the zone of mixing of the California Cur-
rent with the Equatorial Water mass of the eastern tropi-
cal Pacific. Hence, through recent time, most tropical
euphausiid species and those of the southern part of the
California Current have had access to waters of the Gulf.
Present-day colonization by species from the California
Current system has been made by only three species, rep-
resenting three genera, whereas tropical species appear as
transients, except in the southernmost part of the Gulf.
Nematoscelis difJicilis, Nyctiphanes simplex,
and
Euphausia eximia
are established in much of the Gulf,
and their ranges there varied little with season.
Euphau-
sia pacifica
and
Thysanoessa gregaria
are distributed
like
N. diflcilis
in the California Current, and both may
occur as far south as the mouth of the Gulf (Brinton
1979).
Their absence in the Gulf is evidently due to
inability to accommodate to summer extremes in temper-
ature and to a lack of flow into the Gulf in winter-spring
when conditions there might be within their ranges of
tolerance.
E. pacipca
is a vertical migrator which enters
the surface layer at night in all regions where the species
has been observed. This characteristic could be a dis-
advantage for residence in the Gulf.
N. diflcilis
tends to
avoid the surface layers where a thermocline is devel-
oped, as in the summer in the Gulf.
Thysanoessa gre-
garia
is a nonmigrator or short-distance migrator inhabit-
ing the depth range of the thermocline itself. Evidently
such a stratum is insufficiently stable or constant in
temperature to have permitted
T.
gregaria
to occur here.
Nyctiphanes simplex
and
Euphausia eximia
are logi-
cal inhabitants of the Gulf because their ranges are cen-
tered in these latitudes. In addition, the extensive shelf in
the Gulf is particularly suitable for
N. simplex.
Many euphausiid species occupy different depths, both
in the course of their life histories and diurnally as mi-
grating juveniles and adults. These depths may span
hundreds of meters. Distributions of these species are
certainly affected by the conservative conditions beneath
the sea surface. However, the generally more variable
upper level phenomena-currents, temperature, food-
may act in a critical way at any stage of an animal's
development. Variation in some, if not all,
of
these para-
meters is more extreme in the Gulf, particularly the
northern half, than in most oceanic regions. The August
warming to
29-3
1
"C in the
Gulf
is more extreme than in
the open ocean, and the
Gulf
euphausiids then enter a rest-
ing phase with respect to reproduction, though the ranges
of most species are then most expanded.
The biogeographical accommodations that euphausi-
ids in the Gulf of California have made to seasonal
changes in temperature and direction of flow include both
reciprocal and common responses among the warm-tem-
perate and the tropical species in this partly closed sys-
tem. Reciprocal responses include the relationships of the
ranges of
Nematoscelis dflcilis
and
N.
gracilis
in
the
232

BRINTON AND TOWNSEND: EUPHAUSIIDS
IN
THE GULF OF CALIFORNIA-THE 1957 CRUISES
CalCOFI Rep., Vol.
XXI,
1980
bo
Figure
23.
The tropical euphasiids (four species: Euphausia distinguenda,
E.
lamelligera, E. tenera,
and
Nematoscelis gracilis) shown
as
%
Of
total
euphausiids occuring in the
Gulf
of
California
southern half of the Gulf during February-April, with
tropical
N. gracilis
occupying areas where warm-tem-
perate
N. dzzcilis
was absent or at low density and mov-
ing northward on the.eastern side in June as
N. dzzcilis
receded from there. Both species of
Nematoscelis
showed
lowest density and diminished range during the heat of
August (Figures 8, 9).
Although the range of
Euphausia eximia
extended
northward into mid-Gulf during all seasons, the composite
range of the strictly tropical species showed northern
range extension after April, reaching the zone of the island
channels in June and the northern Gulf in August where
their dominance extended northward through mid-Gulf
(Figure
23).
The dominant Gulf species,
Nyctiphanes simplex
and
Nematoscelis dzzcilis,
although both occupying most of
the Gulf during all seasons, nevertheless showed certain
complementary characteristics.
N. dzzcilis
was at its
peak
in
mid-Gulf in February, while
N. simplex
was then
building for April (western Gulf) and June (northern
Gulf) maximum abundances. By June
N. diflcilis
was
sparse in the northern Gulf (Figures 7,8). In August,
N.
simplex
was most dense in the zone of the island chan-
nels, where
N. dflcilis
was, for the first time, sparse
or absent.2
'An unpublished report by Mundhenke
(1969:
The relationships between water masses and
euphausiids in the GulfofCalifornia and the eastern tropical Pacific, masters thesis,
U.S.
Naval
Postgraduate School.
I
I5
pager) has come
to
our attention since this paper went
to
press.
This
discusses euphausiids irom
54
TuckerTrawl sampler obtained by
RfV
Te Vega
intheCulfduring
September 22-November
14.1961
As
in
our August samples,
Euphausia exximia
and
E. distin-
Though there were distinct zonal irregularities in the
presence of larvae (Figure 18), there were trends showing
that
N. simplex
calyptopes and, to a lesser extent, fur-
cilia: were more abundant along the shallower eastern
side of the Gulf during their periods of peak production,
whereas maxima for
N. dzzcilis
were in the generally
deeper waters of the western side (Figure 20B). However,
the shallow northern
Gulf
was an important nursery area
for both species.
The strong warm-temperate zooplankton commun-
ity in the Gulf includes, together with the euphausiids
Nematoscelis dzzcilis
and
Nyctiphanes simplex,
the
copepod
Calanus
paczpcus californicus
Brodskii which
bulked as the dominant organism in the Gulf during
February-April.
Moser et al. (1 974) emphasized the widespread occur-
rence in the Gulf of two fishes of commercial importance
having cold-water affinities. The Pacific mackerel,
Scom-
ber japonicus,
averaged fifth in abundance in the Gulf,
and the Pacific sardine,
Sardinops sagax caeruleus,
var-
ied between third and sixth in abundance, depending
on
the cruise.
According to A. Fleminger (personal communication),
most
Calanus
submerge to greater depths than the 0-1 40m
sampled during these 1957 cruises. Evidence that this
is not the case with
N. dflcilis
consists of the fact that
guenda
occurred northward to the island channels but
hemoroscelisgracrlrs
was
In
mid
Gulf
21
N
as
in
ourJune samples
E lamelligera E reitem
and the
Strlocheirons
were
not
recorded
but
E gibboides
and
Nemorobrachronfleripes
were reported inside the Gulfto
28
N
diffenng
significantly from our findings
Nemaroscelis dtflcils
was found
rn
all zones but
N~criphanes
simplex
only from mid
Gulf
northward
233

BNNTON AND TOWNSEND: EUPHAUSIIDS
IN
THE GULF
OF
CALIFORNIA-THE 1957 CRUISES
CalCOFI Rep.,
Vol.
XXI,
1980
numbers of adult
N. diflcilis
did not decrease signifi-
cantly in August in the 0-140 m layer sampled during the
1957 cruises (Figure 17). Younger stages were thenmuch
reduced, but this is consistent with the pattern in all
species.
The values in the Gulf for biomass of euphausiids were
25 cc/l,OOO m3 during February to June, dropping to 7 cc
in
August. The higher means are like those in the richer
part of the California Current off central California, San
Francisco to Point Conception (Isaacs et al. 1969; Flem-
inger et al. 1974). There, 30-80 cc/l,OOO m3 were mea-
sured along different lines of stations between San Fran-
cisco and Point Conception during April 1956. Mean
values of biomass for different east-west transects of the
California Current were then 4-9 cc from Point Concep-
tion southward along Baja California. During April in the
warm year 1958, values were 3-6 cc/l,OOO m3 all the way
from San Francisco southward.
Summer (July) values for euphausiid biomass during
1955-56 intheCaliforniacurrent were 10-15 cc/1,000m3
north of Point Conception and 3-10 cc to the south. Values
in 1957-59 were slightly lower. The mean proportion
of total biomass made up by euphausiids was also near-
ly the same on opposite sides of the Baja California
peninsula.
In
the southern part of the California Current
during 1956-59 it was 10% (January), 7% (April), 6%
(July), 10% (October), comparing with 7% (February),
5% (April), 7% (June) and 3% (August) in the Gulf. The
richer places along western Baja California for both eu-
phausiids and total biomass are near Punta Banda (3 1 ON),
Vizcaino Bay (28-29"N), and Punta Eugenia (29"N)
southward along the coast. In the eastern tropical Pacific
(Brinton 1979), euphausiids were at maxima of 13 cc and
26 cc beneath 1 m2, or 20% and
50%
oftotal volume, at the
equator and the northern edge of the North Equatorial
Countercurrent. Off the mouth of the Gulf they were 2-7
cc/l,OOO m3, or 3-6% of total biomass. The values for cc
beneath 1 m2 approximately convert to cc per 1,000 m3
by doubling. Longhurst's (1976) data indicated that eu-
phausiids comprise about 5% of total biomass at 14"N in
the eastern tropical Pacific, agreeing -&ith proportions
observed at the mouth of the Gulf.
Thus, euphausiid biomass and its proportion of the
total biomass in the Gulf -is intermediate between that
found in the richest and poorest parts of either the Cali-
fornia Current or the eastern tropical Pacific. Parts of the
Gulf having the largest standing stocks correspond to
places and times where
Nyctiphanes simplex
and
Nema-
toscelis diflcilis
are most abundant. At such localities,
values of 50-200 cc/l,OOO m3 for euphausiids and
>
1,000 cc for total biomass are as high as any reported
from mid- to low latitudes in the Pacific.
Could
Nyctiphanes simplex
be
economically harvested
in the Gulf? Several localities having densities of
>5O,OOO/
1,000m3 were encountered in the northern Gulf during
April-June. Such densities of euphausiids are rare in the
California Current, although
N. simplex
has been found
in such numbers in Bahia Sebastian Vizcaino, 28"N. In
the northern Gulf, the mean biomass of this species was at
a maximum of 77 g/l,OOO m3 in the western sector
in
June (data converted from Figure 18A). This is some-
what higher than April and June values of 35 g and 48 g
from the eastern sector of the northern Gulf.
As stated in the section on euphausiid biomass, a
factor of 2 may be applied to these values for
N. simplex
from CalCOFI 1-m net catches so as to equate them with
those obtained using an unbridled net.
This
gives mean
biomass values in the range of 70-1 50 g/l,OOO m3 in the
sectors in which
N.
simplex
is most abundant. Higher
local densities would, of course, be encountered.
The range 70-150 g compares with 100-300 g/l,OOO
m3 for
Euphausia pacifica
in the vicinity of the Strait of
Georgia in the inland waterway of British Columbia.
There,
E. pacipca
and
Calanusplumchrus
have been the
objects of a modest fishery, which expanded from 10 tons/
year in 1970 to
>lo0
tons in 1974 (Heath 1977). Mid-
water trawls were used.
A next step in addressing the question of practicability
of harvesting
N. simplex
would be exploratory fishing in
the northern Gulf by means of plankton trawls, possibly
in conjunction with acoustic reconnaissance (e.g. Sam-
eoto 1972), and a study of population dynamics of the
species.
LITERATURE CITED
Ahlstrom,
E.H.
1948. A record of pilchard eggs and larvae collected
during surveys made in 1939 to 1941.
U.S.
Fish. Wildl. Serv., Spec.
Sci. Rep. Fish. 54, 75 p.
Alvariiio, A. 1969. Zoogeografia del Mar de Cortes: Quetognatos,
Siphonoforos y Medusas.
An.
Inst. Biol. Univ. Nal. Autbn. Mexico
40. Ser. Cienc. del Mar
y
Lmnol. 1:ll-54.
Anonymous.
1963. CalCOFI atlas of 10-metertemperatures and salin-
ities 1949 through 1959. Calif. Coop. Oceanic Fish. Invest. Atlas 1,
ii-v, charts.
Ayala-Castaiiares, A. 1979. Palabras del Doctor Agustin Ayala-
Castaiiares Coordinador de la Investigacion Cientifica, en la Cere-
monia de Inauguracion de
10s
Seis Simposios Cientificos. Secretaria
de Rectoria, Direccion General de Divulgacion, Universidad Na-
cional Autbnoma de Mexico, 29 de Enero de 1979.
Boden, B.P. 195 1. The egg and larval stages of
Nyctkhanes
simplex,
a euphausiid crustacean from California. Proc. Zool. SOC. Lond.,
121 (pt. II):515-527.
Bradshaw,
J.S.
1959. Ecology
of
living planktonic foraminifera
in
the
north and equatorial Pacific Ocean. Cushing Found. Foram. Res.,
Contrib., 10:25-64.
Brinton,
E. 1962. The distribution of Pacific euphausiids. Bull. Scripps
Inst. Oceanogr. 8(2):51-270.
234

BFUNTON AND TOWNSEND: EUPHAUSIIDS
IN
THE GULF OF CALIFORNIA-THE 1957 CRUISES
CalCOFI Rep.,
Vol.
XXI,
1980
.
1967a. Vertical migration and avoidance capability of
euphausiids in the California Current. Limnol. Oceanogr. I2(3):45 1-
483.
. 1967b. Distributional atlas of Euphausiacea (Crustacea)
in the California Current region, Part
I,
Calif. Coop. Oceanic Fish.
Invest. Atlas 5, iii-xi, charts 1-275.
. 1973. Distributional atlas of Euphausiacea (Crustacea)
in the California Current recion, Part
11,
Calif. Coop. Oceanic Fish.
Invest. Atlas 18, v-vii, charts 1-336.
.
1975. Euphausiids of Southeast Asian waters. Naga Re-
ports, Scripps Inst. Oceanogr., Univ. Calif., 4(5) 287 p.
. 1976. Population biology of
Euphausia paci$ca
off
southern California. Fish. Bull. U.S. 74(4):733-762.
.
1979. Parameters relating to the distribution of planktonic
organisms, especially euphausiids in the eastern tropical Pacitic.
Prog. Oceanogr.
8(
3):
125-1 89.
Brinton, E., D. Siegel-Causey, and A. Fleminger.
(in
press) Zoe
plankton in the Gulf of California. Proc. Six Sci. Sym. IV. The Gulf
of California: origin, evolution, waters, marine life and resources.
Brinton,
E.,
and J.G. Wyllie. 1976. Distributional atlas of euphausiid
growth
stages
off
southern California, 1953 through 1956. Calif.
Coop. Oceanic Fish Invest. Atlas 24,
vii-xxxii.
charts 1-289.
Ekman,
S.
1953. Zoogeography of the sea. Sidgwick and Jackson,
London, 417 p.
Emilsson,
I.,
and M.A. Alatorre, (in press) Recent investigations on the
circulation in the outer part of the Gulf of California. Proc. Six
Sci. Sym. IV. The Gulf of California: origin, evolution, waters,
marine life and resources.
Fleminger, A. 1967. Taxonomy, distribution and polymorphism in the
Labidocera jollae
group with remarks on evolution within the group
(Copepoda: Calanoida). Proc.
U.S.
Nat. Mus. 120(3567):1-61.
.
1975. Geographical distribution and morphological diver-
gence in American coastal-zone planktonic copepods of the genus
Labidocera.
P. 392-419
In
Estuarine research, Vol.
I,
Chemistry,
biology and the estuarine system. Academic Press, New York.
Fleminger, A,, J.D. Isaacs, and J.G. Wyllie. 1974. Zooplankton bic-
mass measurements from CalCOFI cruises of July 1955 to 1969 and
remarks on comparison with results from October, January, and
April cruises of 1955 to 1959. Calif. Coop. Oceanic Fish. Invest.
Atlas 21, vi-xx, charts 1-1 18.
Gilbert, J.Y., and W.E. Allen. 1943. The phytoplankton of the Gulf of
California obtained by the E.W. Scripps in 1939 and 1940. J. Mar.
Res. 5(2):89-I
10.
Gilmartin, M.. and N. Revelante. 1978. The phytoplankton character-
istics of the barrier island lagoons of the Gulf of California. Estuar.
Coastal Mar. Sci. 7(1):29-47.
Gopalakrishnan, K. 1973. Developmental and growth studies of the
euphausiid
Nematoscelis dificilis
(Crustacea) based on rearing.
Bull.
Scripps Inst. Oceanogr., Univ. Calif. 20-1-87.
Gros, P., and J. Cochard. 1978. On the biology
ofNyctiphanes couchii
(Crustacea: Euphasusiacea) in the northern Bay of Biscay. Ann.
Inst. Oceanogr., Paris, 54( 1):25-46.
Heath, W.A. 1977. The ecology and harvesting of euphausiids in the
Strait of Georgia. Ph.D. Thesis. Univ. British Columbia. 187 p.
Isaacs, J.D., A. Fleminger, and J.K. Miller. 1969. Distributional atlas
of
zooplankton biomass in the California Current region: Spring and
Fall 1955-1959. Calif. Coop. Oceanic Fish. Invest. Atlas 10, v-xxv,
charts 1-252.
Jorgensen,
G.,
and J.B.L. Matthews. 1975. Ecological studies on the
deepwater pelagic community
of
Korsfjorden, Western Noway.
Population dynamics
of
six species of euphausiids in 1968 and 1969.
Sarsia 59:67-84.
LeRoux, A. 1973. Observations sur le developpement larvaire de
N.vcriphanes couchii
(Crustacea: Euphausiacea) au laboratoire. Mar.
Biol. 22:159-166.
Longhurst, A.R. 1976. Interaction between zooplankton and phytc-
plankton profiles in the eastern tropical Pacific Ocean. DeegSea
Res. 23:729-754.
Love. C. (ed.) 1973. EASTROPAC Atlas. National Oceanic and
Atmospheric Administration; Fish. Circ. 330, 7, 1-8 and figs.
Mauchline, J., and L.R. Fisher. 1969. The biology of euphausiids.
Adv. Mar. Biol. 7. Academic Press, London
&
New York, 454 p.
Moser, H.G., E.H. Ahlstrom, D. Kramer, and E.G. Stevens. 1974.
Distribution and abundance of fish eggs and larvae
in
the Gulf of
California. Calif. Coop. Oceanic Fish. Invest. Rep. 17:l 12.130.
Newman, W.A. 1979. California transition zone: Significance of short-
range endemics. P. 399-416
In
J. Gray and A.J. Boucot (eds.),
Historical biogeography, plate tectonics. and the changing environ-
ment. Oregon State Univ. Press.
OsorieTafal, B.F. 1946. Contribucion a1 conocimiento del Mar de
Cortes. Bol. de la SOC. Mex. de Geografia
y
Estadistica, 62( 1):
89-130.
Parker, F.L. 1973. Living planktonic foraminifera from the Gulf of
California. J. Foram. Res. 3(2):70-77.
Reid. J.L.. Jr.. R.S. Arthur. and E.B. Bennett (eds.) 1965. Oceanic
observations of the Pacific: 1957. Univ. Calif. Press, Berkeley and
Los Angeles, 707 p.
Roden, G.I. 1971. Large-scale upwelling
off
northwestern Mexico. J.
Phys. Oceanogr. 2: 184- 189.
Roden.
G.1.,
and G.W. Groves. 1959. Recent oceanographic investiga-
tions in the Gulf of California. J. Mar. Res. 18(
1):10-35.
Round, F.E. 1967. The phytoplankton of the Gulf of California. Part I.
Its composition, distribution and contribution to the sediments. J.
Exp. Mar. Biol. Ecol. 1(1):76-97.
Sameoto, D.D. 1972. The feasibility of mapping euphausiid concentra-
tions
in
the Gulf of St. Lawrence using high frequency sonar. J. Fish.
Res. Board Can., Tech. Rep. No. 350, 30 p.
Siegel-Causey, D. (in press) Pelagic amphipoda of the Gulf of Cali-
fornia. Proc. Six Sci. Symp. IV. The Gulf of California: origin,
evolution, waters, marine life and resources.
Steinbeck, J., and E.F. Ricketts. 1941. The sea of Cortez. Viking
Press, New York, 598 p.
Walker, B.W. 1960. The distribution and affinities of the marine fish
fauna of the Gulf of California.
In
Symposium: the biogeography of
Baja California and adjacent seas. Pt. 2 Marine biotas. Zyst. Zool.
9(3-4):123-133.
Wiebe, P.H., and
S.H.
Boyd. 1978. Limits of
Nemaroscelis megalops
in the northwestern Atlantic in relation to Gulf Stream cold core
rings. I. Horizontal and vertical distributions. J. Mar. Res. 36(1):
119.142.
Wiebe. P.H., and
L.
D'Abramo. 1972. Distribution
of
euphausiid
assemblages in the Mediterranean Sea. Marine Biol.
15:
130-1
49.
Wyllie, J.G. 1966. Geostrophic flow of the California Current at the
surface and at 200 meters. Calif. Coop. Oceanic Fish. Invest. Atlas
4, vii-xiii, charts 1-288.
Zeitzschel, B. 1969. Primary productivity in the Gulf of California.
Mar. Biol. 3(3):201-217.
235

BRINTON AND TOWNSEND: EUPHAUSIIDS IN THE GULF OF CALIFORNIA-THE
1957
CRUISES
CalCOFI Rep.,
Vol.
XXI,
1980
00;;
0000
oom-
oos0
00-u
w
*
mu
m
%
2
V
.-
c
W
2
I*
oopm
0000
oomo
oowo
0032
N
-
c
?
m
3
3
d
0
4-
0
k
3
c3
oomr-
oo'oo
0-r-w
0000
OONVIo-mo
UN
"NN
0
mN
u0
2
n
m
L
3
0
LL
e,
5
E
t
0
m
e,
0
e,
Q.
v)
.-
3
m
r
3
a
W
z
0
m
L
2
m
u
5
*
cr
d
3
cr
m
W
L
I/
2
d
B
I1
v'
I*
s
.-
E!
5
c
L?
B
I1
?
-
X
z
w
p.
-
n
a
a
236
